















-
CRISPR Knockout KitCRISPR Point Mutation KitKI Enhancer Drug
-
Precision Mutation Cell PanelsKnock-in cell lineWild Type Cell Line
-
-
 Gene Expression & Regulation - Positive Regulation of pri-miRNA Transcription
Gene Expression & Regulation - Positive Regulation of pri-miRNA Transcription
MicroRNAs (miRNAs) are small non-coding RNAs that broadly regulate gene expression at the post-transcriptional level. Their primary transcripts (pri-miRNAs) are synthesized by RNA polymerase II (Pol II). Transcription of pri-miRNAs is tightly regulated by multiple transcription factors, coactivators, and signaling pathways, and dysregulation is closely associated with cancer, cardiovascular diseases, developmental disorders, and metabolic diseases.
Gene knockout cell lines targeting key regulators of this pathway enable investigation of transcriptional regulatory networks controlling miRNA expression, identification of tissue-specific miRNA regulatory elements, and therapeutic target discovery for related diseases. EDITGENE has established a ready-to-use KO cell line library covering more than 60 validated targets involved in pri-miRNA transcription regulation, including SRF, MYC, FOS, JUN, STAT3, TP53, SMAD3/4, HIF1A, PPARG, AR, EGFR, and related genes to support non-coding RNA and transcription regulation research.
Transcription of pri-miRNAs is primarily mediated by RNA polymerase II. Promoter regions of pri-miRNAs typically contain multiple transcription factor binding sites, and positive regulators activate transcription of specific miRNA clusters by responding to extracellular signals—including growth factors, cytokines, hormones, hypoxia, TGF-β/BMP, and Wnt/Notch signaling—or through direct interaction with the Pol II complex.
Core Positive Regulators Include:
· Serum Response Factor (SRF) and Coactivators:SRF forms complexes with MRTFA, MRTFB, and MYOCD to regulate smooth muscle- and cardiac-enriched miRNAs such as miR-1, miR-133, and miR-145.
· SMAD Transcription Factors:SMAD1, SMAD3, and SMAD4 mediate TGF-β/BMP signaling (via upstream BMP2, BMPR1A, TGFB1/2) and induce fibrosis- and epithelial-mesenchymal transition-related miRNAs, while SMAD6 acts as a feedback regulator.
· STAT3 and Inflammation-Related Factors:STAT3 responds to IL6 and IL10 signaling and cooperates with RELA (NF-κB subunit), FOS/JUN (AP-1), and FOSL1 to regulate immune- and inflammation-associated miRNAs.
· Tumor-Associated Transcription Factors:TP53 activates the miR-34 family; MYC and MYCN broadly promote transcription of miRNA clusters such as miR-17~92; ETS1 regulates angiogenesis-related miRNAs; and WT1 controls kidney development-associated miRNAs.
· Hypoxia and Metabolic Regulators:HIF1A induces miR-210 under hypoxic conditions; PPARG regulates adipogenesis-associated miRNAs such as miR-143; SREBF1/2 control lipid metabolism-related miRNAs; and NR1H2 (LXRβ) participates in cholesterol homeostasis.
· Hormone Signaling Regulators:AR responds to androgen signaling and, together with FOXA1, GATA2, and GATA3, regulates prostate-specific miRNAs; PRL and NGFR participate in neuroendocrine-associated miRNA regulation; and NR3C1 regulates stress-responsive miRNAs.
· Development and Stem Cell Factors:PAX6, POU2F1 (Oct1), NOTCH2, NOTCH3, KLF4, EGR1, and ATOH8 regulate miRNA expression during embryonic development, neurogenesis, and stem cell maintenance.
· Chromatin Remodeling and Telomere Factors:SMARCA4 (BRG1) promotes Pol II recruitment through chromatin remodeling, while TERT and GNL3 participate in nucleolar structure and coupling of miRNA transcription.
· Autocrine and Paracrine Signaling Molecules:FGF2, PDGFB, EGFR, TNF, APLN, and AGT indirectly promote pri-miRNA transcription by activating downstream transcription factors through their respective receptors.
· RNA Metabolism-Associated Regulators:GTPBP2, HBS1L, CSDE1, PELO, CNOT6, and TESK1 mainly function in RNA metabolism but may positively regulate pri-miRNA levels by influencing transcription elongation or transcription-degradation coupling mechanisms.
Loss of function of these genes can reduce expression of specific miRNA clusters, thereby affecting cell proliferation, differentiation, apoptosis, and immune responses, contributing to diseases such as cancer, cardiac hypertrophy, pulmonary fibrosis, and diabetic nephropathy.
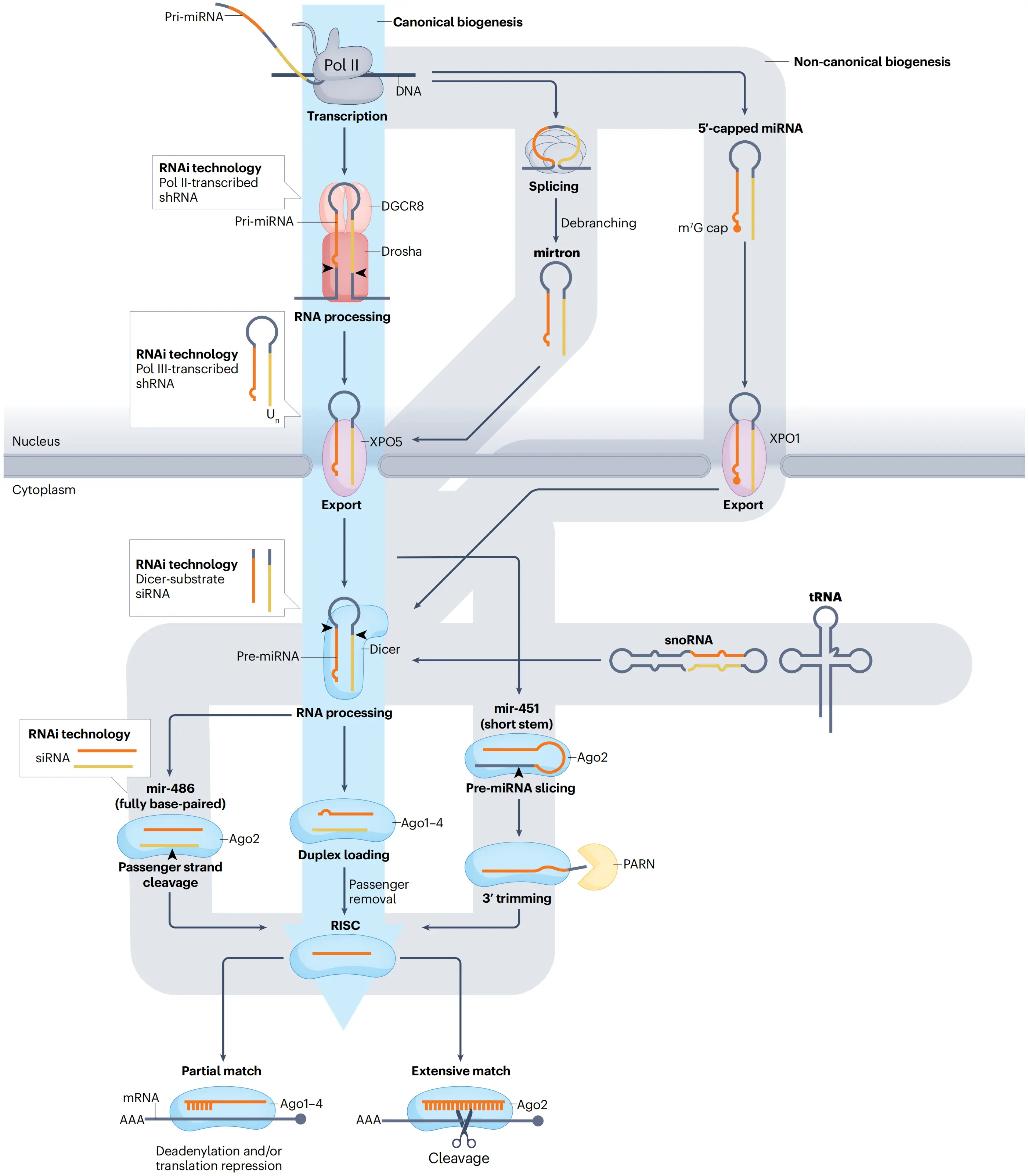
The microRNA biogenesis pathway and its use in RNAi technology
(Kim et al., Nat Rev Mol Cell Biol, 2016)
Abnormal positive regulation of pri-miRNA transcription is associated with multiple diseases, including cancers (such as hepatocellular carcinoma, breast cancer, and prostate cancer), cardiac hypertrophy and heart failure, pulmonary hypertension, pulmonary fibrosis, renal fibrosis, diabetic vascular complications, psoriasis, neurodevelopmental disorders, and lipid metabolism disorders.
Gene knockout cell models provide ideal tools for investigating how transcription factors regulate specific pri-miRNAs and how signaling networks shape miRNA expression programs, supporting mechanism studies, target validation, and drug discovery.
· Tumorigenesis and Metastasis Models
Used to investigate the effects of TP53, MYC, FOS, JUN, STAT3, ETS1, or WT1 knockout on oncogenic or tumor-suppressive miRNAs such as miR-34, miR-21, and miR-155, and evaluate transcription factor inhibitors including STAT3 or MYC inhibitors.
· Cardiovascular and Fibrosis Disease Models
Used to study the effects of SRF, MRTF, MYOCD, SMAD3, SMAD4, TGFB1, or NOTCH3 knockout on cardiac/smooth muscle-enriched miRNAs and fibrosis-associated miRNAs, supporting screening of anti-hypertrophic and anti-fibrotic therapeutics.
· Metabolic Disease and Hormone Signaling Models
Used to investigate the effects of PPARG, SREBF1/2, HIF1A, AR, FOXA1, GATA2/3, or NR1H2 knockout on adipogenesis-, cholesterol metabolism-, and hormone-responsive miRNAs.
· Development and Stem Cell Models
Used to study the roles of PAX6, NOTCH2, KLF4, EGR1, POU2F1, or ATOH8 knockout in miRNA transcriptional regulation in embryonic stem cells or neural progenitor cells.
· RNA Metabolism Coordination Models
Used to investigate whether GTPBP2, HBS1L, CSDE1, PELO, CNOT6, or TESK1 knockout indirectly alters pri-miRNA levels through effects on transcription elongation or RNA stability.
EDITGENE’s positive regulation of pri-miRNA transcription (RNA Pol II) knockout cell line library includes validated ready-to-use models covering more than 60 key regulatory factors involved in the pathway. These models support research on miRNA transcriptional regulation, signaling pathways and miRNA networks, and disease-associated transcription factor function.
In addition, we provide both ready-to-use and customized knockout cell line services to support research in cancer, cardiovascular disease, development, and metabolic disorders. Additional targets and cell types—including HEK293, A549, HepG2, H9c2, and RAW264.7—are available upon request.
-
Cat.No: EDC90437
Species: Human
Cell Name: HEK293
Gene Name: DRD2
Gene ID: 1813
Specs: 1×10⁶cells
-
Cat.No: EDC00205
Species: Human
Cell Name: hTERT-RPE1
Gene Name: TP53
Gene ID: 7157
Specs: 1×10⁶cells
-
Cat.No: EDC10177
Species: Human
Cell Name: HeLa
Gene Name: GTPBP2
Gene ID: 54676
Specs: 1×10⁶cells
-
Cat.No: EDC07921
Species: Human
Cell Name: HaCaT
Gene Name: HIF1A
Gene ID: 3091
Specs: 1×10⁶cells
-
Cat.No: EDC08198
Species: Human
Cell Name: HAP1
Gene Name: JUN
Gene ID: 3725
Specs: 1×10⁶cells
-
Cat.No: EDC08315
Species: Human
Cell Name: HEK293T
Gene Name: HIF1A
Gene ID: 3091
Specs: 1×10⁶cells
-
Cat.No: EDC09412
Species: Human
Cell Name: HAP1
Gene Name: TESK1
Gene ID: 7016
Specs: 1×10⁶cells
-
Cat.No: EDC90244
Species: Human
Cell Name: BEAS-2B
Gene Name: PPARG
Gene ID: 5468
Specs: 1×10⁶cells
-
Cat.No: EDC90542
Species: Human
Cell Name: NALM6, clone G5
Gene Name: TP53
Gene ID: 7157
Specs: 1×10⁶cells
-
Cat.No: EDC07854
Species: Human
Cell Name: HCT 116
Gene Name: TP53
Gene ID: 7157
Specs: 1×10⁶cells
-
Cat.No: EDJ-KQ110
Species: Human
Cell Name: HEK293T
Gene Name: NR1H2
Gene ID: 7376
Specs: 1×10⁶cells
-
Cat.No: EDJ-KQ126
Species: Human
Cell Name: HEK293
Gene Name: SMAD6
Gene ID: 4091
Specs: 1×10⁶cells
-
Cat.No: EDJ-KQ176
Species: Human
Cell Name: HEK293
Gene Name: JUN
Gene ID: 3725
Specs: 1×10⁶cells
-
Cat.No: EDJ-KQ184
Species: Human
Cell Name: HEK293T
Gene Name: JUN
Gene ID: 3725
Specs: 1×10⁶cells
-
Cat.No: EDJ-KQ210
Species: Human
Cell Name: HEK293
Gene Name: NGFR
Gene ID: 4804
Specs: 1×10⁶cells
- 1
- 2
- ...
- 16
- 17
- Next Page »
Subscribe
You can unsubscribe from these communications at any time. For more information on how to unsubscribe, our privacy practices, and how we are committed to protecting and respecting your privacy, please review our Privacy Policy.
By clicking submit below, you consent to allow EDITGENE to store and process the personal information submitted above to provide you the content requested.
